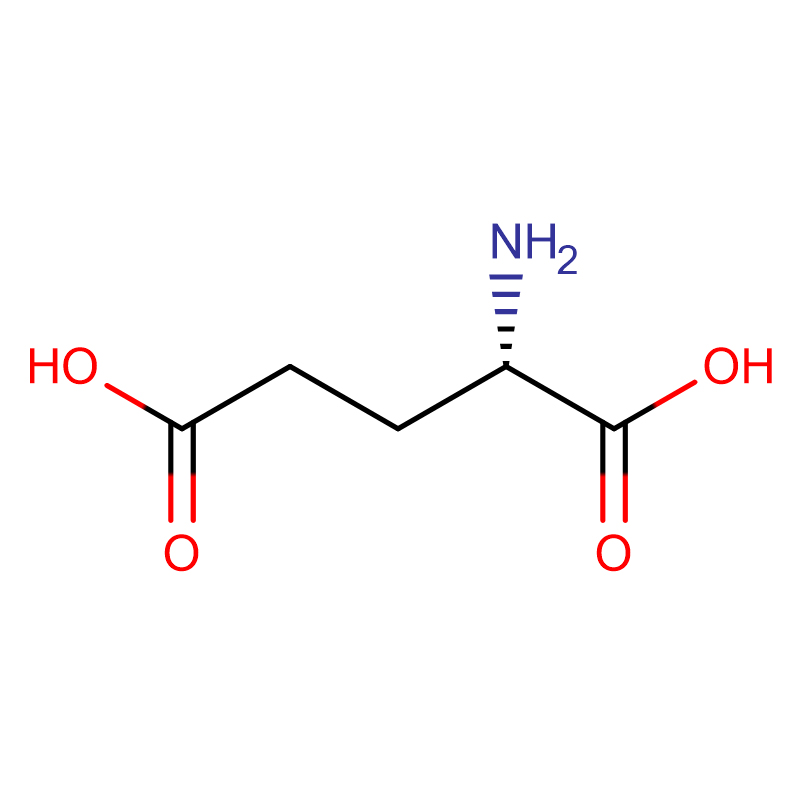
L-glutaminska kiselina CAS:56-86-0 99% bijeli prah
| Kataloški broj | XD90312 |
| ime proizvoda | D-glutaminska kiselina |
| CAS | 56-86-0 |
| Molekularna formula | C5H9NO4 |
| Molekularna težina | 147.13 |
| Pojedinosti o pohrani | Ambijentalni |
| Harmonizirani tarifni broj | 29224200 |
Specifikacija proizvoda
| Izgled | Bijeli kristal ili kristalni prah |
| Ispitivanje | 99% |
| Specifična rotacija | +31,5 do +32,5° |
| pH | 3,0 do 3,5 |
| Gubitak sušenjem | 0,2% maks |
| Željezo | 10 ppm max |
| AS2O3 | 1 ppm max |
| Teški metal (Pb) | 10 ppm max |
| Amonij | 0,02% maks |
| Ostale aminokiseline | <0,4% |
| Klorid | 0,02% maks. |
| Ostatak nakon žarenja (sulfatiziran) | 0,1% maks |
| Sulfat (kao SO4) | 0,02% maks |
EAAT2 prijenosnik glutamata, odgovoran je za >90% hipokampalnog unosa glutamata.Iako se EAAT2 pretežno eksprimira u astrocitima, oko 10% molekula EAAT2 nalazi se u terminalima aksona.Unatoč nižoj razini ekspresije EAAT2 u glutamatergičkim terminalima, kada se rezovi hipokampusa inkubiraju s niskom koncentracijom d-aspartata (supstrat EAAT2), terminali aksona nakupljaju d-aspartat jednako brzo kao i astroglija.To implicira neobjašnjivu neusklađenost između distribucije proteina EAAT2 i transportne aktivnosti posredovane EAAT2.Jedna je hipoteza da je (1) heterorazmjena unutarnjeg supstrata s vanjskim supstratom znatno brža od neto unosa i (2) terminali favoriziraju heterorazmjenu zbog visokih razina unutarnjeg glutamata.Međutim, trenutačno je nepoznato imaju li heterorazmjena i preuzimanje slične ili različite stope.Kako bismo riješili ovaj problem, koristili smo rekonstituirani sustav za usporedbu relativnih stopa dvaju procesa kod štakora i miševa.Neto unos bio je osjetljiv na promjene u membranskom potencijalu i bio je stimuliran vanjskim propusnim anionima u skladu s postojanjem nesprežene anionske vodljivosti.Koristeći potonje, također pokazujemo da brzina heterorazmjene također ovisi o membranskom potencijalu.Osim toga, naši podaci dalje ukazuju na prisutnost curenja natrija u EAAT2.Uključivanjem novih otkrića u naš prethodni model preuzimanja glutamata pomoću EAAT2, predviđamo da je voltažna osjetljivost izmjene uzrokovana trećim vezanjem Na(+) ovisno o voltaži.Nadalje, naši eksperimenti i simulacije sugeriraju da su relativne stope neto preuzimanja i heterorazmjene usporedive u EAAT2.